Medicle Datenbank: Eine kurze Einführung in die Biochemie der viralen Infektion
Medicle Datenbank: Eine kurze Einführung in die Biochemie der viralen Infektion |
| Aufbau von Viren | ||||
| Aufbau von Viren | ||||
|
I. Aufbau von Viren Das Virus-Genom kann aus RNA- oder DNA aufgebaut sein, wonach dementsprechend di Viren bezeichnet werden: als RNA-Viren oder als DNA-Viren. Geimeinsam ist beiden, daß sie eine Proteinhülle besitzen, das Capsid. Diese Hülle ist aus einzelnen Capsomeren, Untereinheiten des Capsids, aufgebaut. Morphologisch lassen sich dabei Ikosaeder von unregelmäßigen helikalen Strukturen (helikale Nucleocapsidsymmetrie) unterscheiden. Zu diesem Capsid besitzen viele Viren noch eine weitere Hüllmembran, deren Lipide meist aus der Membran der Wirtszelle stammen. | ||||
|
Funktionell ist die Nucleinsäure der Träger der Informationen über den Aufbau und die Wirkung des Virus, die Proteinhülle bestimmt die Gestalt und die Antigenität, schützt den Virus vor nucleinsäureabbauenden Enzymen und bestimmt die Wechselwirkungen des Virus mit der Zielzelle und der Membranoberfläche. | ||||
|
Da eine Virus-Zielzellen-Interaktion über Rezeptoren zustande kommt, kann man davon ausgehen, daß Viren durch eine Kreuzreaktion mit einem Rezeptor für ein körpereigenes Molekül agieren, weil Zellen wohl keinen Rezeptor für eine Virus-Interaktion besitzen. | ||||
|
Viren lassen sich nach ihrem Genom unterscheiden, was sehr in seiner Größe variieren kann (von 3200 Basenpaaren bis zu 250.000-300.000 Basenpaaren). Bei sehr kleinen Viren wird oftmals durch eine Leserasterverschiebung eine optimale Ausnutzung des Genoms bewirkt. Dadurch können verschiedene Elemente des Virus „übereinander“ codiert werden. Die Nucleinsäure codiert für Strukturproteine (die meist als Polyproteine synthetisiert werden und durch limitierte Proteolyse entstehen), für Enzymproteine (die für die Replikation und die Integration in das Wirtszellengenom notwendig sind) und für Regulatorproteine (die die Transkription des Virusgenoms beeinflussen). | ||||
| RNA-Viren | ||||
|
II. RNA-Viren Die RNA eines RNA-Virus läßt sich in zwei Kategorien einteilen: in Virus-Genom mit positiver Polarität oder mit negativer Polarität. Die Polarität ergibt daraus, ob die RNA direkt als Matrize für die Proteinbiosynthese verwendet werden kann. Bei positiver Polarität bedeutet das: die RNA ist aus Sequenzen aufgebaut, die der zellulären mRNA entsprechen. | ||||
|
Bei negativer Polarität bedeutet das: die RNA ist aus Sequenzen aufgbaut, die komplementär zur mRNA sind. Doppelsträngige RNA-Viren besitzen positive und negative RNA, die zusammen die genomische RNA bilden. Bei positiver Polarität des Virus-Genoms wird die Virus-RNA zur ribosomalen Proteinsynthese herangezogen und die dabei entstehende viruscodierte RNA-abhängige RNA-Polymerase zur Replikation des Virus-Genoms verwendet. | ||||
|
Bei negativer Polarität (bzw. Doppelstrang) ergibt sich eine besondere Situation, da die genomische RNA erst durch ein viruscodiertes Enzym transkribiert werden muß. Diese Gruppe der RNA-Viren heißt Retroviren. Das viruscodierte Enzym ist die reverse Transkriptase, welches RNA in DNA umschreibt (um dann in das Wirtsgenom integriert zu werden). Das Genom von Retroviren codiert für: ein Strukturprotein (das mit der RNA im Kern des Virus assoziiert ist: das gruppenspezifische Antigen oder GAG-Protein), eine Polymerase (POL-Protein) und ein Glykoprotein (das sich in der Virusproteinhülle befindet und die Bindung des Virus an die Wirtszellenmembran bewirkt: ENV-Protein, von engl. „enevelope“ Hülle). Dieses Bauprinzip ist bei allen Retroviren gleich. | ||||
|
Zur Nomenklatur: Onkornaviren = onkogene RNA Viren, Picornaviren = pico RNA Viren (kleinste RNA-Viren). | ||||
| HIV-1 | ||||
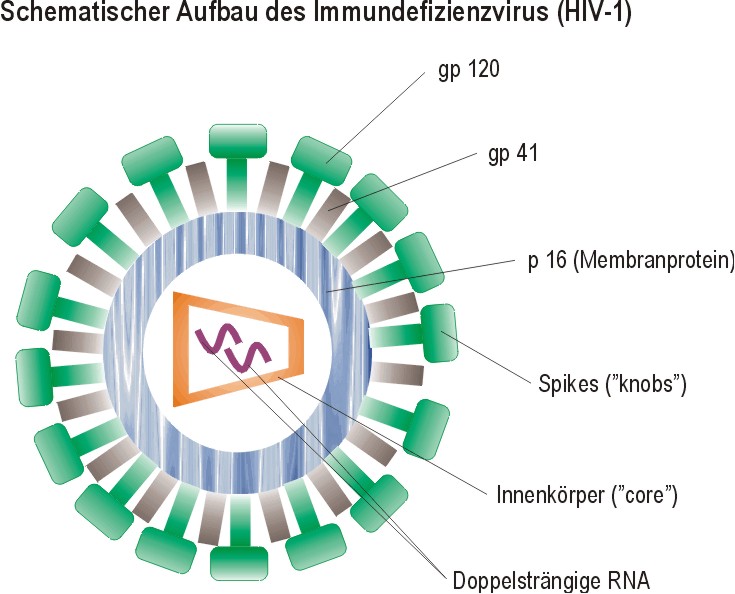
III. Humanes Immundefizienz Virus (HIV-1) Das HIV-1 ist ein doppelstängiges Retrovirus, dessen Capsid die Struktur eines Kegelstumpfes besitzt, die Hüllmembran besteht aus 72 ENV-Proteinen. Das Virus hat in seinem Genom zehn Gene, was bedeutet, daß es neben den GAG-, POL- und ENV-Proteinen noch für weitere Proteine codiert. Das sind die sogennanten LTR-Gene (long terminal repeats), die am 5´- und am 3´-Ende des Genoms liegen und zwei Funktionen haben: die effiziente Transkription des Virusgenoms (durch Promotor / Enhancer-Funktionen) und die Förderung der Integration der gebildeten Virus-DNA in das Genom. | ||||
|
Das GAG-Gen codiert für das Polyprotein p53 (d.h. Molekulargewicht 53 kDa), das mit Myristinsäure verestert ist. Diese langkettige Fettsäure erlaubt die Wechselwirkung des Proteins mit Lipiden der Hüllmembran. Aus dem GAG-Polyprotein entstehen durch limitierte Proteolyse die Capsid-Proteine p17 und p24, sowie die im Viruskern befindlichen p15, p7 und p9. | ||||
|
Die Transkription des POL-Gens führt zur Bildung einer p150-Vorstufe, aus der durch limitierte Proteolyse drei Enzyme entstehen: p16, eine Protease, die das GAG-Polyprtotein spaltet, p51, die reverse Transkriptase, p34 eine DNA-Endonuklease oder -integrase, die ander Integration der gebildeten Virus-DNA in das Wirtszellengenom beteiligt ist. Die limitierte Proteolyse der Polyproteine geschieht durch die HIV-1-Protease, die durch HIV-1-Protease-Inhibitoren in der AIDS-Therapie gehemmt wird. Das ENV-Gen codiert für das Glykoprotein gp160, das ein wichtiger Bestandteil der Hüllmembran des Virus ist. Aus Spaltung dieses Proteins geht ein membranständiges Protein gp 41 und ein freies Protein gp 120 hervor, wobei beide nicht covalent miteinander verknüpft sind! Dadurch geht dem Virus beständig gp 120 verloren. Zusätzlich enthält das HIV-Genom noch weitere Regulatorgene, deren Funktion noch nicht geklärt ist. | ||||
|
Das HIV-1 bindet über das Glykoprotein 120 seiner Hüllmembran an das CD-4-Protein (das sich auf T4-Lymphozyten, epidermalen Langerhans-Zellen und Monozyten bzw. Makrophagen befindet und normalerweise MHC-Klasse-II Moleküle bindet). Durch Interaktion mit gp 41 und unter Mitwirkung des Membranproteins Fusin wird das Nucleocapsid in die Zelle gebracht, wo dann die Auflösung der Capsidmembran und die Freisetzung der Nucleinsäuren erfolgt. | ||||
|
Nun folgt die Eklipse, eine Phase, in der die Virusteilchen nicht mehr zu erkennen sind. Nach Herstellung von cDNA (die RNA dient als Matrize) durch die virale reverse Transkriptase, wird die nun doppelsträngige DNA unter der Einwirkung des Enzyms Integrase als Provirus-DNA in das Wirtsgenom eingebaut. Durch Bindung des zellulären Transkriptionsfaktors NF-kB (an die LTR-Regionen des Virusgenoms) wird die Transkription der HIV-1-DNA gestartet. Durch regulatorische Proteine wird die Tranksription vertausendfacht. Danach wird die virale mRNA synthetisiert und nach Transfer ins Zytosol durch Translation zur Bildung von Virusstruktur- und Virusenzym-Proteinen verwendet. | ||||
|
Die synthetisierten ENV-Proteine des virus gelangen auch die Membran des Wirtes, wo sie auf der Oberfläche exprimiert werden und auf der Innenseite einen „Rezeptor“ für die neu assoziierenden viralen „Bausteine“ darstellen. Das halbfertige Nucleocapsid tritt nun an diese Membran heran, koppelt mit den ENV-Proteinen und wird nach außen gestülpt und abgeschnürt (was dazu führt, daß die umhüllende Membran aus viralen und aus wirtseigenen Proteinen aufgebaut ist). | ||||
|
Da Viren keinen Reparatur-Apparat für ihr Genom haben, kann es auch zu Mutationen innerhalb der reversen Tranksiptase kommen. Das bewirkt eine schnelle Entstehung von Virusmutanten, die es schwierig machen, die HIV-Infektion zu behandeln. | ||||
| Adenoviren | ||||
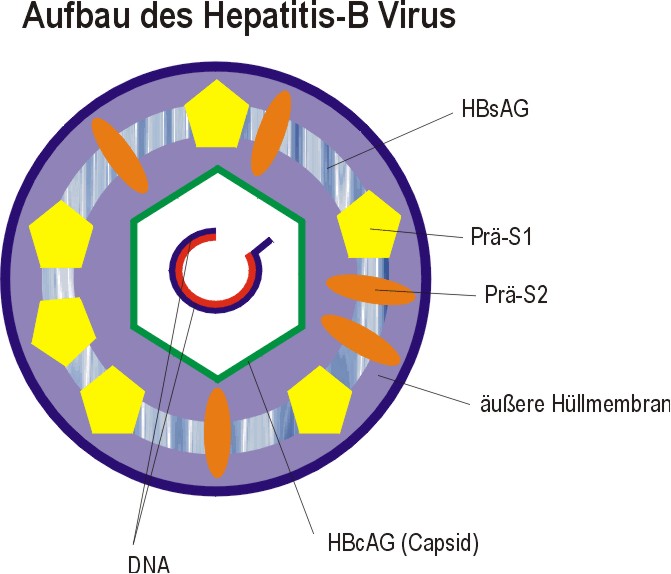
IV. Hepatitis-B Virus Das Hepatitis-B Virus-Genom ist extrem klein und codiert durch Leserasterverschiebung für mehrere Proteine. Dabei enthält die äußere Hüllmembran drei strukturell verwandte Proteine (neben den Lipiden der Wirtszelle), die alle eine Domäne von 226 Aminosäuren besitzen. Diese Proteine sind in kleines S-Protein, ein mittleres Prä-S2-Protein, ein großes Prä-S1-Protein | ||||
|
Beide Prä-Proteine ragen aus der Hüllmembran heraus und bestimmen so die Interaktion mit anderen Zellen. Das Capsid (die innere Kapsel) wird nur von dem HBc-Antigen gebildet, einer einzigen Proteinart, die die Virusnucleinsäure umgibt. Dabei ist der Plusstrang um 20-50% kürzer als der gegenläufige Minusstrang. Durch die Ringstruktur bildet sich durch Überlappung aus. Die codierten Gene sind:
| ||||
|
Die Replikation des HB-Virus-Genomes geschieht in mehreren Schritten. Nach dem das Virus in die Wirtszelle aufgenommen wurde, wird die Hüllmembran abgestreift und das Capsid in den Zellkern transportiert. Dort wird die offene ringförmige Virus-DNA in eine zirkuläre DNA verwandelt, um anschließend von den zelleigenen RNA-Polymerasen abgelesen und in RNA transkribiert zu werden. | ||||
|
Dies ist ein ungewöhnlicher Weg für DNA-Viren, normalerweise geschieht die Replikation direkt, ohne die RNA-Zwischenstufe! Die virale DNA wird nicht in das Wirtszellengenom integriert (da diese Viren keine Integrase als Enzym besitzen), sie verbleibt aber im Zellkern und ist somit in vielen Zellen des Körpers ein „Reservoir“ an Virus-DNA. | ||||
|
Diese neuentstandenen Ribonucleinsäuren codieren für die viralen Proteine (vier virale mRNAs) und für ein sogenanntes Prä-Genom, das zusammen mit der viralen Polymerase in ein neugebildetes Capsid gepackt wird und ins Cytosol transportiert wird. Dort schreibt die virale Polymerase das RNA-Prä-Genom in einen DNA-Minusstrang um, was der Funktion der reversen Transkriptase von Retroviren entspricht. Anschließend wird die RNA-Vorlage von der RNAse H zerstört und der komplementäre DNA-Strang wird synthetisiert. Dadurch entsteht ein offenes ringförmiges Molekül. Dieser letzte Schritt der Replikation der Virus-DNA findet bereits während der Ausschleusung des Virus aus der Wirtszelle statt, so dass die Synthese des Plus-Stranges mit dem Austreten der Zelle stoppt und damit diese Region eine variable Länge erhält. | ||||
|
Eine hohe Mutationsrate (auch durch den komplizierten Syntheseweg über die RNA-Zwischenstufe) gibt diesen Viren einen Selektionsvorteil, da das Immunsystem gegen immer neuen Varianten ankämpfen muss. | ||||
| Hepatitis-B Virus | ||||
V. Adenoviren Diese Virenart verursacht beim Menschen lokale Infektionen, besonders der Atemwege und des Gastrointestinaltraktes. Es handelt sich dabei um doppelsträngige DNA-Viren mit einer Ikosaederstruktur. Sie besitzen einen Durchmesser von 60-90nm und haben keine Hüllmembran. Adenoviren infizieren eine Vielzahl von Zellen, was darauf schließen lässt, dass ein ubiquitärer Membranrezeptor noch nicht identifiziert worden ist. Die darauf folgende Internalisierung erfolgt über eine Integrinbindung mit der Pentonbasis. Über die sogenannten coated pits wird es dann ins Cytosol aufgenommen. Dort wird dann durch die Ruptur der Phagolysosomenmembran das Nucleocapsid in das Cytosol abgegeben. Nach Auflösung des Capsids wird die virale Nucleinsäure in den Zellkern der Wirtszelle transferiert. Diese Phase wird auch als Eklipsephase bezeichnet, da in dieser Zeit das Virus aus der Zelle „verschwunden“ scheint. Die Genexpression des Virusgenoms beginnt ca. 8h nach Infektion und ist abhängig von der Expression des E1-Genprodukts. Das bedeutet für den therapeutischen Einsatz als Vektoren eine gute Kontrolle über die Replikationsfähigkeiten des Adenovirus.Die späteren Genprodukte sind die Capsidproteine: Hexon, Penton und Faserproteine (Aufbau: siehe Schema rechts). Die Hexone (orange in der Darstellung rechts) bedecken die Kanten und Oberflächen (die aus mehreren gleichseitigen Dreiecken aufgebaut ist), während die Pentone mit den Faserproteinen (beide grün in der Darstellung rechts) die „Antennen“ an den Scheitelpunkten bilden. Da die meisten Adenoviren die Wirtseigene Transkription und Translation hemmen kommet es nach einiger Zeit zu einem lytischen Prozeß, der die Wirtszelle zerstört (lytische Infektion). | ||||
|
Neben den Adenoviren existieren noch sogennante adenoassoziierte Viren. Solche Viren gehören zu der Gruppe der Parvoviren und sind beim Menschen nicht pathogen, sie sind aus einzelsträngiger DNA aufgebaut. Ihre Bedeutung liegt deshalb besonders in der Gentherapie. Um ihren Replikationszyklus durchführen zu können, benötigen diese Viren einen Helferadeno- oder Herpesvirus. Fehlt diese Koinfektion, dann wird zwar das adenoassoziierte Virusgenom in das Wirtsgenom integriert aber nicht repliziert. | ||||
| Referenzen | ||||
Lehrbuch |
||||
 |
||||
| ||||
| Editorial | ||||
Autor |
||||
|
||||
Erstellt |
03.12.2002 |
|||
|
||||
Freigabe |
ALL |
|||
|
||||
Status |
TRACK3 |
|||
|
||||
Copycheck |
1 - überprüft |
|||
| ||||
| Kommentare | ||||