Medicle Datenbank: Eine kurze Einführung in die Biochemie der viralen Infektion
Medicle Datenbank: Eine kurze Einführung in die Biochemie der viralen Infektion |
| Adenoviren | ||||
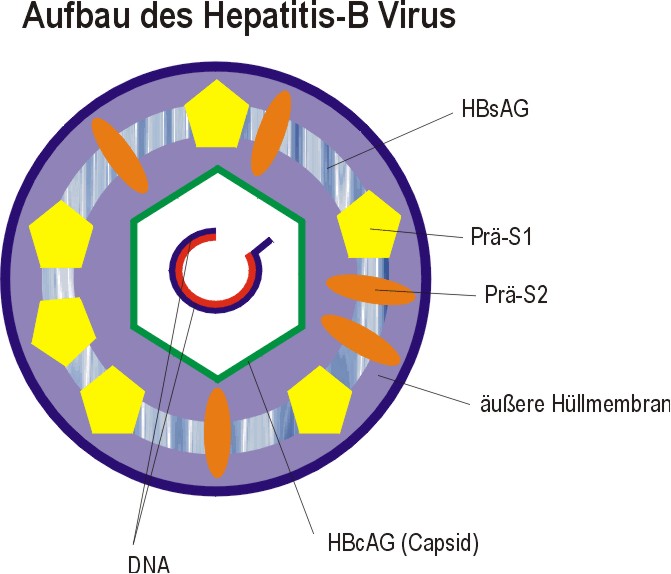
IV. Hepatitis-B Virus Das Hepatitis-B Virus-Genom ist extrem klein und codiert durch Leserasterverschiebung für mehrere Proteine. Dabei enthält die äußere Hüllmembran drei strukturell verwandte Proteine (neben den Lipiden der Wirtszelle), die alle eine Domäne von 226 Aminosäuren besitzen. Diese Proteine sind in kleines S-Protein, ein mittleres Prä-S2-Protein, ein großes Prä-S1-Protein | ||||
|
Beide Prä-Proteine ragen aus der Hüllmembran heraus und bestimmen so die Interaktion mit anderen Zellen. Das Capsid (die innere Kapsel) wird nur von dem HBc-Antigen gebildet, einer einzigen Proteinart, die die Virusnucleinsäure umgibt. Dabei ist der Plusstrang um 20-50% kürzer als der gegenläufige Minusstrang. Durch die Ringstruktur bildet sich durch Überlappung aus. Die codierten Gene sind:
| ||||
|
Die Replikation des HB-Virus-Genomes geschieht in mehreren Schritten. Nach dem das Virus in die Wirtszelle aufgenommen wurde, wird die Hüllmembran abgestreift und das Capsid in den Zellkern transportiert. Dort wird die offene ringförmige Virus-DNA in eine zirkuläre DNA verwandelt, um anschließend von den zelleigenen RNA-Polymerasen abgelesen und in RNA transkribiert zu werden. | ||||
|
Dies ist ein ungewöhnlicher Weg für DNA-Viren, normalerweise geschieht die Replikation direkt, ohne die RNA-Zwischenstufe! Die virale DNA wird nicht in das Wirtszellengenom integriert (da diese Viren keine Integrase als Enzym besitzen), sie verbleibt aber im Zellkern und ist somit in vielen Zellen des Körpers ein „Reservoir“ an Virus-DNA. | ||||
|
Diese neuentstandenen Ribonucleinsäuren codieren für die viralen Proteine (vier virale mRNAs) und für ein sogenanntes Prä-Genom, das zusammen mit der viralen Polymerase in ein neugebildetes Capsid gepackt wird und ins Cytosol transportiert wird. Dort schreibt die virale Polymerase das RNA-Prä-Genom in einen DNA-Minusstrang um, was der Funktion der reversen Transkriptase von Retroviren entspricht. Anschließend wird die RNA-Vorlage von der RNAse H zerstört und der komplementäre DNA-Strang wird synthetisiert. Dadurch entsteht ein offenes ringförmiges Molekül. Dieser letzte Schritt der Replikation der Virus-DNA findet bereits während der Ausschleusung des Virus aus der Wirtszelle statt, so dass die Synthese des Plus-Stranges mit dem Austreten der Zelle stoppt und damit diese Region eine variable Länge erhält. | ||||
|
Eine hohe Mutationsrate (auch durch den komplizierten Syntheseweg über die RNA-Zwischenstufe) gibt diesen Viren einen Selektionsvorteil, da das Immunsystem gegen immer neuen Varianten ankämpfen muss. | ||||