Medicle Datenbank: Eine kurze Einführung in die Biochemie der viralen Infektion
Medicle Datenbank: Eine kurze Einführung in die Biochemie der viralen Infektion |
| HIV-1 | ||||
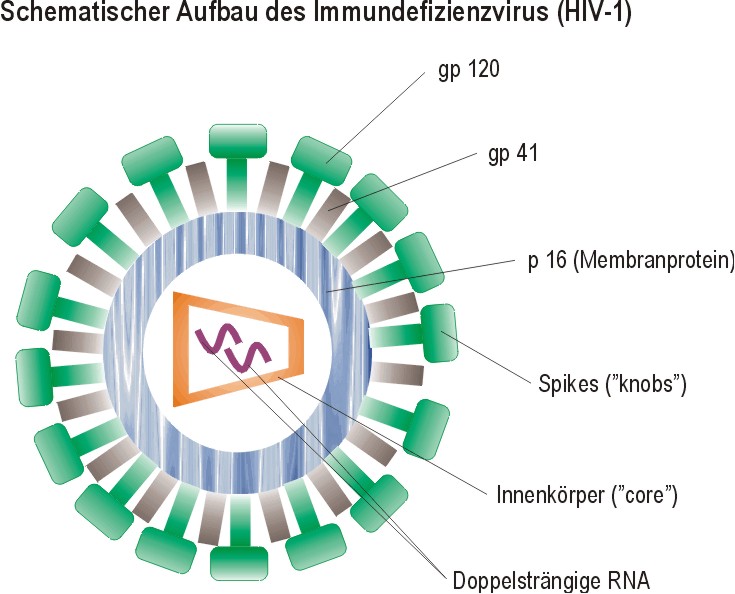
III. Humanes Immundefizienz Virus (HIV-1) Das HIV-1 ist ein doppelstängiges Retrovirus, dessen Capsid die Struktur eines Kegelstumpfes besitzt, die Hüllmembran besteht aus 72 ENV-Proteinen. Das Virus hat in seinem Genom zehn Gene, was bedeutet, daß es neben den GAG-, POL- und ENV-Proteinen noch für weitere Proteine codiert. Das sind die sogennanten LTR-Gene (long terminal repeats), die am 5´- und am 3´-Ende des Genoms liegen und zwei Funktionen haben: die effiziente Transkription des Virusgenoms (durch Promotor / Enhancer-Funktionen) und die Förderung der Integration der gebildeten Virus-DNA in das Genom. | ||||
|
Das GAG-Gen codiert für das Polyprotein p53 (d.h. Molekulargewicht 53 kDa), das mit Myristinsäure verestert ist. Diese langkettige Fettsäure erlaubt die Wechselwirkung des Proteins mit Lipiden der Hüllmembran. Aus dem GAG-Polyprotein entstehen durch limitierte Proteolyse die Capsid-Proteine p17 und p24, sowie die im Viruskern befindlichen p15, p7 und p9. | ||||
|
Die Transkription des POL-Gens führt zur Bildung einer p150-Vorstufe, aus der durch limitierte Proteolyse drei Enzyme entstehen: p16, eine Protease, die das GAG-Polyprtotein spaltet, p51, die reverse Transkriptase, p34 eine DNA-Endonuklease oder -integrase, die ander Integration der gebildeten Virus-DNA in das Wirtszellengenom beteiligt ist. Die limitierte Proteolyse der Polyproteine geschieht durch die HIV-1-Protease, die durch HIV-1-Protease-Inhibitoren in der AIDS-Therapie gehemmt wird. Das ENV-Gen codiert für das Glykoprotein gp160, das ein wichtiger Bestandteil der Hüllmembran des Virus ist. Aus Spaltung dieses Proteins geht ein membranständiges Protein gp 41 und ein freies Protein gp 120 hervor, wobei beide nicht covalent miteinander verknüpft sind! Dadurch geht dem Virus beständig gp 120 verloren. Zusätzlich enthält das HIV-Genom noch weitere Regulatorgene, deren Funktion noch nicht geklärt ist. | ||||
|
Das HIV-1 bindet über das Glykoprotein 120 seiner Hüllmembran an das CD-4-Protein (das sich auf T4-Lymphozyten, epidermalen Langerhans-Zellen und Monozyten bzw. Makrophagen befindet und normalerweise MHC-Klasse-II Moleküle bindet). Durch Interaktion mit gp 41 und unter Mitwirkung des Membranproteins Fusin wird das Nucleocapsid in die Zelle gebracht, wo dann die Auflösung der Capsidmembran und die Freisetzung der Nucleinsäuren erfolgt. | ||||
|
Nun folgt die Eklipse, eine Phase, in der die Virusteilchen nicht mehr zu erkennen sind. Nach Herstellung von cDNA (die RNA dient als Matrize) durch die virale reverse Transkriptase, wird die nun doppelsträngige DNA unter der Einwirkung des Enzyms Integrase als Provirus-DNA in das Wirtsgenom eingebaut. Durch Bindung des zellulären Transkriptionsfaktors NF-kB (an die LTR-Regionen des Virusgenoms) wird die Transkription der HIV-1-DNA gestartet. Durch regulatorische Proteine wird die Tranksription vertausendfacht. Danach wird die virale mRNA synthetisiert und nach Transfer ins Zytosol durch Translation zur Bildung von Virusstruktur- und Virusenzym-Proteinen verwendet. | ||||
|
Die synthetisierten ENV-Proteine des virus gelangen auch die Membran des Wirtes, wo sie auf der Oberfläche exprimiert werden und auf der Innenseite einen „Rezeptor“ für die neu assoziierenden viralen „Bausteine“ darstellen. Das halbfertige Nucleocapsid tritt nun an diese Membran heran, koppelt mit den ENV-Proteinen und wird nach außen gestülpt und abgeschnürt (was dazu führt, daß die umhüllende Membran aus viralen und aus wirtseigenen Proteinen aufgebaut ist). | ||||
|
Da Viren keinen Reparatur-Apparat für ihr Genom haben, kann es auch zu Mutationen innerhalb der reversen Tranksiptase kommen. Das bewirkt eine schnelle Entstehung von Virusmutanten, die es schwierig machen, die HIV-Infektion zu behandeln. | ||||